
Agradecimentos ao amigo Johnny pelo desenho autoral, para conhecer o trabalho de divulgação dele no youtube, que tem tudo a ver com o nosso, clique aqui.
No primeiro texto da série tratamos da questão histórica que envolve o problema da organização da diversidade biológica. Também definimos um conceito fundamental para tentar resolver objetivamente a questão de como organizar os seres vivos: o conceito de grupo natural. Para entender essa definição, leia a primeira parte aqui.
Na segunda parte, nos aprofundamos em algumas ideias de Hennig, fundador da Sistemática Filogenética, e construímos o conceito de sinapomorfia, os atributos que servem de base e sustentação dos grupo naturais a partir de alguns critérios de homologia. A parte 2, disponível aqui, é fundamental para compreender este texto, e a sugestão é que ele seja lido antes de prosseguirmos.
Lendo um cladograma
Vamos inicialmente ver como se lê um cladograma, que é a representação gráfica das relações de parentesco entre os seres vivos. Saber ler um cladograma é um passo fundamental para entendermos o exercício. Observe a figura abaixo:

Um cladograma apresenta algumas informações essenciais, como por exemplo as relações entre táxons ou relação de grupos-irmãos. C e D são grupos-irmãos em relação a B, o que significa que o último ancestral comum entre C e D (ancestral z) é mais recente do que o último ancestral compartilhado entre B e D, por exemplo (ancestral y); isso sugere que há uma relação evolutiva mais íntima entre C e D do que entre B e D ou entre C e B.
É importante entendermos que o relevante nessa representação é a relação de grupos-irmãos, e não a posição dos grupos. Por exemplo os dois cladogramas abaixo são idênticos do ponto de vista filogenético:

Em ambos, C é grupo irmão de D em relação a B, e CDB formam um grupo que exclui A. Apenas o posicionamento gráfico se altera, mas as relações de parentesco permanecem as mesmas.
Agora vamos à um exercício que vai, além de revisar os conceitos do texto 2, ilustrar na prática quais os passos para irmos da observação de organismos e chegarmos aos tão procurados grupos naturais organizados em árvores.
Antes de iniciarmos, é sempre bom deixar claro que esse é apenas um exercício para apresentar a lógica por trás do método cladístico. Tanto os grupos de interesse quanto os caracteres foram selecionados de forma didática. Na prática, as coisas são bem mais complicadas que isso; árvores gigantescas são geradas a partir de matrizes com dezenas e as vezes centenas de caracteres.
Passo 1 - Definindo o grupo de interesse
Quando estava pensando em qual grupo de organismos seria interessante para fazermos esse exercício, cheguei a três conclusões.
A primeira é de que ele não pode ser muito abrangente, pois seria pouco didático tentar desenvolver relações de homologia entre estruturas bastante modificadas, mesmo que sejam derivadas de uma estrutura ancestral comum. A segunda, em contraponto, é de que não poderia ser um grupo muito restrito, como ao nível de ordem, pois além de ainda haver muitos conflitos filogenéticos nesse nível, muitas estruturas específicas teriam de ser usadas (o que demandaria inúmeros parênteses no texto, muito mais do que os que eu já uso normalmente). Por último, cheguei a conclusão que o grupo deveria ser familiar à todos, pois além de evitar múltiplas explicações adicionais (e muitos parênteses) é sempre mais interessante para o leitor que tratemos de coisas com as quais ele está familiarizado.
O grupo escolhido foi o dos tetrápodes, que compreende todas as linhagens de vertebrados terrestres: anfíbios, répteis, aves e mamíferos.
Passo 2 - Escolha de táxons representativos
Em sistemática, os terminais, que podem ser outros grupos ou espécies (como no nosso caso) são chamados de táxons. Selecionei 7 animais como táxons representativos e genéricos para a construção dos nossos grupos:
-Sapo
-Pombo
-Crocodilo
-Humano
-Cobra
-Ornitorrinco
-Morcego
Passo 3 - Grupo externo
Esse passo é muito importante, pois um grupo externo é o que nos dá como base quais características seriam externas ao grupo e portanto, ao menos em hipótese, plesiomórficas em relação aos tetrapoda. Esse processo se chama polarização, e o recomendado é que se use o grupo mais aparentado possível do seu grupo de interesse, para evitar um excesso de convergências entre o grupo externo e os internos. No nosso caso, o grupo considerado irmão dos tetrapoda são os peixes pulmonados (Dipnoi).

Pirambóia (Lepidosiren paradoxa), uma das 6 espécies vivas de peixes sarcopterígios pulmonados, encontrada em bacias de água doce da América do Sul, inclusive na Amazônica. Imagem disponível aqui.[1]
Passo 4 - Hipóteses de homologia primárias
Para sugerir caracteres, ou seja hipóteses de homologia, os três critérios de homologia já citados no segundo texto foram usados. Forma, posição e ontogenia (para mais detalhes, veja o segundo texto da série).
A partir desses 3 critérios foram levantados os seguintes caracteres para os tetrápodes:
1 - 4 membros diferenciados, sustentando o corpo
2 - Embrião menos dependente de água (com Ovo Amniótico)*
3 - Glândulas Mamárias
4 - Placenta
5 - Asas
6 - Forame mandibular**
7 - Glândulas mucosas na pele
8 - Duas fenestras temporais***
2 - Embrião menos dependente de água (com Ovo Amniótico)*
3 - Glândulas Mamárias
4 - Placenta
5 - Asas
6 - Forame mandibular**
7 - Glândulas mucosas na pele
8 - Duas fenestras temporais***
A importância evolutiva dessas características para a evolução dos tetrápodes foge do escopo deste texto e será no futuro (olha o spolier) tratado separadamente aqui no blog.
*Ovo Amniótico: no caso dos mamíferos com placenta, esse ovo amniótico é internalizado e modificado em placenta, além de alguns anexos ,como o saco vitelínico, estarem atrofiados já que a nutrição fica por conta da mãe através do cordão umbilical.
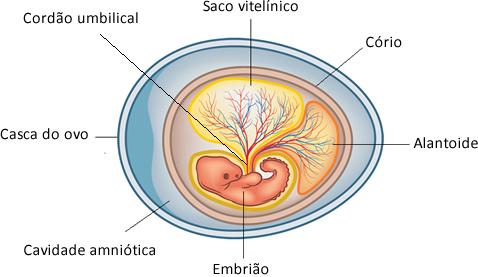
Imagem disponível aqui.
**Forame mandibular:
 |
| Adicionar legenda |
Imagens mostrando as aberturas mandibulares no crânio de um pombo e um crocodilo. Imagens aqui e aqui.
***Duas fenestras temporais no crânio:

Crânio com duas fenestras temporais, imagem disponível aqui.
Passo 5 - Construção da matriz
Agora que o levantamento das características foi feito, vamos colocá-las em uma matriz com a a seguinte estrutura:
Linhas = táxons
Coluna = hipóteses de homologia

Preenchemos a matriz com 0 para ausência da característica e 1 para presença.

OBS: Esse não é a única forma de preencher a matriz, elas permitem caracteres com mais de dois estados (multiestado), porém esse preenchimento binário chamado de A/P (ausente-presente) é o mais simples e intuitivo.
Passo 6 - Teste de congruência
Quando organismos compartilham um mesmo estado de caráter (o 1 na nossa matriz), significa que há uma hipótese de agrupamento entre eles.
Ao analisarmos a hipótese 2 (ovo amniótico), ela sugere um grande grupo formado por Crocodilo, Pombo, Cobra, Ornitorrinco, Humano e Morcego. Ou seja, essa característica surge uma vez, no ancestral comum entre todos esses animais, e é herdada pelos seus descendentes. Graficamente teríamos a figura abaixo.


OBS.: Se você está se perguntando o que significa "2(1)", é o seguinte: significa que a característica dois está presente. Lembre-se que escolhemos "1" para indicar presença, e "0" para indicar ausência. De maneira geral, "X(1)" e "X(0)" significam que a característica X está presente ou ausente, respectivamente.
Se olharmos agora para a hipótese 3 (presença de glândulas mamárias), vemos que esta característica é compartilhada por Ornitorrinco, Humano e Morcego. O significado sistemático disso é que este atributo sugere um grupo natural que une os 3 táxons, suportado por essa característica que teria surgido no ancestral comum entre eles.


Agora se sobrepusermos as duas hipóteses, o que teremos é o seguinte:
As hipóteses 2 e 3 além de não entrarem em conflito, acabam melhorando a resolução do cladograma, sendo portanto complementares.

Essa lógica será usada para todos os caracteres, as hipóteses serão comparadas e caso elas sejam congruentes ou pelo menos não excludentes (ou seja, quando tudo dá certo), temos um reforço da hipótese de agrupamento.

Homoplasias: quando nem tudo dá certo
Como exposto no texto 2, casos de surgimento independente ou perda de características ocorrem comumente durante a evolução de todos os grupo. Esses casos complicam as análises, e há um critério especial para quando isso ocorre.
Observe abaixo:
Observe abaixo:
A hipótese 1, que sugere que os outros Tetrapoda formam um grupo natural que exclui as cobras, por conta da ausência de 4 membros diferenciados. Vamos observar uma comparação entre essa hipótese e uma outra que sugere uma redução, uma perda dos membros de maneira posterior. (As convergências estão indicadas pelos retângulos não preenchidos de preto)


Essa ideia ilustra o princípio de parcimônia, um critério utilizado por Hennig para podermos nos decidir entre hipóteses conflitantes. O princípio se baseia na navalha de Occam e diz que, quando hipóteses entram em conflito, devemos preferir aquela mais simples. No caso da sistemática, significa que a melhor hipótese deve ser aquela que exige um menor número de passos evolutivos (surgimento de características) e consequentemente, um menor número de homoplasias [2].
Se voltarmos na comparação anterior, veremos claramente que a hipótese que sugere perda posterior dos membros pelas cobras é muito mais parcimoniosa que aquela que sugere que as cobras nunca possuíram um ancestral com pernas, sendo portanto excluído do grupo dos tetrápodes.
O mesmo pode ser observado na hipótese 5 (presença de asas) que sugere que existe um agrupamento entre morcegos e aves, e que portanto as asas surgiriam uma única vez nos tetrápodes. Porém, ao analisar os cladogramas comparativos abaixo, vemos que esse agrupamento é pouco provável, pois ele exigirá o surgimento de 2 características duas vezes na história evolutiva, além da redução de mais duas, o que seria muito mais improvável que se apenas as asas tivessem surgido independentemente.


O mais parcimonioso portanto seria que, Pombo é táxon irmão de Crocodilo, já Morcego é um táxon irmão de Humanos. Este é o resultado mais parcimonioso para o nosso conjunto de dados. Da mesma forma, as cobras formam um grupo junto com os lagartos, seriam um grupo dentro dos tetrápodes que perdeu os membros.
Assim chegamos a nossa arvore final, ou seja, aquela que melhor explica a evolução dos tetrápodes (nos baseando apenas nas poucas características listadas). Ou seja, chegamos na melhor hipótese de relacionamento evolutivo entre os animais em questão, levando em conta o critério de parcimônia, onde consideramos como a melhor hipótese aquela mais simples, que demanda uma menor quantidade de homoplasias e passos evolutivos.
Assim chegamos a nossa arvore final, ou seja, aquela que melhor explica a evolução dos tetrápodes (nos baseando apenas nas poucas características listadas). Ou seja, chegamos na melhor hipótese de relacionamento evolutivo entre os animais em questão, levando em conta o critério de parcimônia, onde consideramos como a melhor hipótese aquela mais simples, que demanda uma menor quantidade de homoplasias e passos evolutivos.
Grupos, finalmente!
Agora que nossa árvore está montada, podemos identificar os grupos naturais e caracterizar também alguns agrupamentos não-naturais.
Grupos naturais:
Os grupos naturais em sistemática são chamados de grupos monofiléticos Estes grupos devem incluir o ancestral comum hipotético e todos os seus descendentes [3]. Olhando para a nossa árvore podemos ver vários grupos monofiléticos. O grupo dos tetrápodes é monofilético, pois inclui o ancestral comum hipotético (marcado em verde) e todos os seus descendentes. Outro grupo monofilético é o que agrupa Ornitorrincos, Morcegos e Humanos, e inclui seu último ancestral comum (marcado em roxo);este grupo é chamado de Mammalia na literatura científica.


Grupos não-naturais:
Há basicamente 2 tipos de agrupamentos não naturais em sistemática, os grupos parafiléticos e os polifiléticos.
Parafilético: Um grupo que reúne ancestral comum, porém que exclui um ou mais de seus descendentes [3]. Um exemplo de grupo parafilético seria "Répteis" senso comum (cobras, lagartos, tartarugas, crocodilos), pois o grupo das aves também compartilha esse mesmo ancestral, isto é, podemos dizer que aves são répteis. Se incluirmos aves em répteis, aí sim teremos um grupo monofilético.


Polifiléticos: Caso o nosso agrupamento com base nas asas fosse retomado, a partir da nossa nova filogenia, ele seria polifilético, pois agrupa dois terminais, que não compartilham um ancestral comum imediato [3]. Para que haja um grupo natural que inclua pombos e morcegos, ele deve incluir também seu último ancestral comum(marcado em roxo) e todos os descendentes: pombo, morcego, ornitorrinco, humano, cobra, crocodilo. Esse grupo monofilético é chamado na taxonomia atual de Amniota.


Conclusão
Chegamos ao fim de mais um texto sobre organização dos seres vivos. Agora já sabemos na prática qual lógica a sistemática usa para agrupar seres vivos, a partir de hipóteses de homologia, chegando finalmente nos tão procurados grupos naturais, que em sistemática ganham o nome de Grupos Monofiléticos . No próximo texto falaremos um pouco sobre algumas mudanças no método desde que ele começou a ser largamente difundido. Falando sobre a relação entre sistemática e a biologia molecular e análises computacionais.
O texto 4 já saiu você pode acessâ-lo clicando aqui.
Referências
O texto 4 já saiu você pode acessâ-lo clicando aqui.
Referências
|




Nenhum comentário:
Postar um comentário