Clique aqui para ler a parte I
 |
| É isso que um criacionista imagina quando se noticia que tecido mole foi preservado? Imagem: internet. |
Este post é uma tradução. O original, completo, pode ser lido no blog Letters to Creationists. A primeira imagem é uma adição minha.
Mecanismos de preservação de proteína antiga
Schweitzer mudou de foco nos últimos anos, da simples demonstração que fragmentos de proteína sobreviveram por milhões de anos em ossos de dinossauro para a consideração dos mecanismos de esta proteína foi preservada. Em um artigo de 2005 [11], ela reconheceu que "as evidências que apoiam a preservação de biomoléculas endógenas no registro fóssil pré-cenozoico geralmente foram recebida com ceticismo, porque se supõe que as moléculas orgânicas primárias não podem suportar as alterações e quebras que ocorrem durante a diagênese". No entanto, essa opinião cética foi baseada em experimentos em tubo de ensaio que podem não representar adequadamente as condições dentro dos poros do osso de dinossauro:" Experimentos de laboratório projetados para aproximar a diagênese molecular aplicam parâmetros físicos e químicos que normalmente não se encontram na natureza (por exemplo, pH <1, T> 300 ⁰C) e não contabilizam os efeitos protetores da associação mineral. Portanto, sua utilidade como representação de processos diagenéticos a nível molecular em amostras preservadas naturalmente é um pouco limitada." Schweitzer passou a listar 16 estudos anteriores que mostraram que" aminoácidos, peptídeos curtos e amino-acúcares podem persistir dentro de fósseis ao longo de uma ampla distribuição de idade geológica ", mais 14 estudos onde" as técnicas imunológicas identificaram compostos antigênicos em fósseis de diferentes idades e de vários tipos de taxa ".
Schweitzer mudou de foco nos últimos anos, da simples demonstração que fragmentos de proteína sobreviveram por milhões de anos em ossos de dinossauro para a consideração dos mecanismos de esta proteína foi preservada. Em um artigo de 2005 [11], ela reconheceu que "as evidências que apoiam a preservação de biomoléculas endógenas no registro fóssil pré-cenozoico geralmente foram recebida com ceticismo, porque se supõe que as moléculas orgânicas primárias não podem suportar as alterações e quebras que ocorrem durante a diagênese". No entanto, essa opinião cética foi baseada em experimentos em tubo de ensaio que podem não representar adequadamente as condições dentro dos poros do osso de dinossauro:" Experimentos de laboratório projetados para aproximar a diagênese molecular aplicam parâmetros físicos e químicos que normalmente não se encontram na natureza (por exemplo, pH <1, T> 300 ⁰C) e não contabilizam os efeitos protetores da associação mineral. Portanto, sua utilidade como representação de processos diagenéticos a nível molecular em amostras preservadas naturalmente é um pouco limitada." Schweitzer passou a listar 16 estudos anteriores que mostraram que" aminoácidos, peptídeos curtos e amino-acúcares podem persistir dentro de fósseis ao longo de uma ampla distribuição de idade geológica ", mais 14 estudos onde" as técnicas imunológicas identificaram compostos antigênicos em fósseis de diferentes idades e de vários tipos de taxa ".
Schweitzer publicou uma ampla revisão [12] da "Preservação de Tecidos Moles em Vertebrados Mesozoicos Terrestres" em 2011. Citando mais de 200 estudos, ela discutiu muitos casos anteriores de achados de tecido mole entre os fósseis mesozoicos. A era mesozóica é vista como variando de cerca de 252 a 66 milhões de anos atrás, e foi denominada "Idade dos Répteis", uma vez que estes eram os animais dominantes em terra, no ar e no mar. A maioria dos exemplos de preservação de tecidos são da pele e seus apêndices, incluindo escamas, penas e garras. Estes "consistem principalmente em proteínas de queratina duráveis e impermeáveis". A queratina tem alto potencial de preservação "por causa de sua estrutura molecular, sua tendência a formar retículados e seus abundantes aminoácidos não-polares".
No entanto, mesmo esses achados são incomuns, uma vez que a norma é que a pele, bem como o tecido subjacente, se decompõem completamente:
Como o carbono e o nitrogênio que compõem proteínas, DNA, células e tecidos de organismos multicelulares são úteis para micróbios para energia metabólica, os restos orgânicos normalmente são degradados rapidamente após a morte; de fato, em circunstâncias normais, mais de 99% do carbono reduzido que compõe esses componentes é retornado rapidamente ao ciclo do carbono por micróbios. Experimentos tafonômicos mostram que, na maioria dos casos, onde as carcaças inteiras foram depositadas na superfície do solo, elas podem ser completamente esqueletizadas em apenas 2 a 3 semanas e as alterações ligadas à degradação na morfologia/ química celular podem ocorrer em poucos minutos após a morte. Consequentemente, a presença de componentes ou células de tecidos originais em associação com restos fossilizados de organismos extintos mostra que os processos normalmente envolvidos na degradação foram desacelerados ou detidos logo após a morte e antes da deterioração completa.
Como o carbono e o nitrogênio que compõem proteínas, DNA, células e tecidos de organismos multicelulares são úteis para micróbios para energia metabólica, os restos orgânicos normalmente são degradados rapidamente após a morte; de fato, em circunstâncias normais, mais de 99% do carbono reduzido que compõe esses componentes é retornado rapidamente ao ciclo do carbono por micróbios. Experimentos tafonômicos mostram que, na maioria dos casos, onde as carcaças inteiras foram depositadas na superfície do solo, elas podem ser completamente esqueletizadas em apenas 2 a 3 semanas e as alterações ligadas à degradação na morfologia/ química celular podem ocorrer em poucos minutos após a morte. Consequentemente, a presença de componentes ou células de tecidos originais em associação com restos fossilizados de organismos extintos mostra que os processos normalmente envolvidos na degradação foram desacelerados ou detidos logo após a morte e antes da deterioração completa.
A Schweitzer oferece algumas sugestões sobre como essa mitigação da decadência pode surgir. Se uma carcaça de animais seca com bastante rapidez, seus tecidos podem sofrer mudanças que os tornam mais estáveis: "... a dessecação precoce através da mumificação pode tornar estes espécimes alvos principais para a recuperação de biomoléculas além do colágeno".
Pondo de lado, muitas das manchetes atraentes sobre "dinossauros mumificados" são enganosas. Por exemplo, um artigo do National Geographic de 2007 foi subtitulado, " 'Dinossauro Mumificado Encontrado'; Tem Pele Intacta e Tecido ". O que você descobre apenas no meio do artigo é que a pele e outros tecidos foram substituídos por minerais, por isso não é a preservação de tecidos moles orgânicos, mas sim a preservação das formas físicas detalhadas dos tecidos macios originais. Isso é interessante, mas também é razoavelmente bem compreendido, pelo menos em esboço. Lingham-Soliara e Glabb [13] relataram sua análise da microestrutura de colágeno de tecido de golfinhos, python e tartaruga decomposto, que posteriormente foi seco ao ar. Eles descobriram que, apesar desta exposição severa à decomposição e desidratação, "muitas fibras de colágeno mantiveram sua integridade estrutural, mostrando pouca degradação". Além disso, "re-hidratação do tecido desidratado apresentou perda estrutural mínima". Eles concluíram que um caminho viável para a preservação de tecidos macios o suficiente para que ele se torne mineralizado (fossilizado) seria que um animal morra durante um período seco (o que, provavelmente, também matou a maioria dos limpadores), se mumifica e depois se cobre com água ou sedimentos ricos em minerais.
Schweitzer [12] também mencionou estudos anteriores que discutiram a estabilização de tecidos dentro de configurações minerais como os poros de osso:
A estreita associação com a fase mineral (Child 1995) pode atuar de forma semelhante à fixação química (por exemplo, com formaldeído), degradação enzimática e microbiana compensatória (Kharalkar et al., 2009 e referências nele contidas). Isso pode ocorrer porque as enzimas microbianas são muito grandes para a maioria dos poros no osso e porque a fase mineral do osso forma uma barreira à digestão (Trueman & Martill 2002, Turner-Walker 2008). Alternativamente, pode ocorrer porque o tamanho pequeno, a grande área superficial e a reatividade dos cristais minerais ósseos podem inibir a degradação enzimática, em um processo semelhante ao demonstrado pelos grãos de argila (Butterfield, 1990, 2003). Finalmente, as restrições de associação com o mineral podem prevenir o inchaço molecular durante a degradação, impedindo o acesso a locais mais reativos em moléculas (M. J. Collins, comunicação pessoal).
A estreita associação com a fase mineral (Child 1995) pode atuar de forma semelhante à fixação química (por exemplo, com formaldeído), degradação enzimática e microbiana compensatória (Kharalkar et al., 2009 e referências nele contidas). Isso pode ocorrer porque as enzimas microbianas são muito grandes para a maioria dos poros no osso e porque a fase mineral do osso forma uma barreira à digestão (Trueman & Martill 2002, Turner-Walker 2008). Alternativamente, pode ocorrer porque o tamanho pequeno, a grande área superficial e a reatividade dos cristais minerais ósseos podem inibir a degradação enzimática, em um processo semelhante ao demonstrado pelos grãos de argila (Butterfield, 1990, 2003). Finalmente, as restrições de associação com o mineral podem prevenir o inchaço molecular durante a degradação, impedindo o acesso a locais mais reativos em moléculas (M. J. Collins, comunicação pessoal).
Um estudo realizado em 2011 por San Antonio, Schweitzer e outros [14] envolveu a análise de fragmentos (peptídeos) de proteína de colágeno recuperados de ossos de dinossauro e o mapeamento dos resultados em modelos moleculares de colágeno derivados de espécies existentes para determinar a configuração das moléculas de colágeno dentro dos ossos do dinossauro. O resumo, em parte:
Os peptídeos de dinossauro foram localizados em regiões de fibrilas protegidas pela embalagem próxima de moléculas de colágeno e continham poucos aminoácidos ácídicos. Quatro peptídeos foram mapeados para regiões de colágeno cruciais para as interações célula-colágeno e desenvolvimento de tecidos. Os peptídeos de dinossauro não foram representados em partes mais expostas da fibrila de colágeno ou regiões que intermediam a reticulação intermolecular. Assim, as regiões funcionalmente significativas de fibrilas de colágeno que são fisicamente protegidas dentro da fibrila podem ser preferencialmente preservadas em fósseis.
Esses resultados mostram empiricamente que as relações estrutura-função a nível molecular podem contribuir para a preservação seletiva em restos de vertebrados fossilizados ao longo do tempo geológico, sugerem um "motivo de preservação" e reforçam os conceitos atuais que ligam a estrutura do colágeno à função biológica. Esta distribuição não aleatória suporta a hipótese de que os peptídeos são produzidos pelos organismos extintos e sugere um mecanismo químico para a sobrevivência.
Os peptídeos de dinossauro foram localizados em regiões de fibrilas protegidas pela embalagem próxima de moléculas de colágeno e continham poucos aminoácidos ácídicos. Quatro peptídeos foram mapeados para regiões de colágeno cruciais para as interações célula-colágeno e desenvolvimento de tecidos. Os peptídeos de dinossauro não foram representados em partes mais expostas da fibrila de colágeno ou regiões que intermediam a reticulação intermolecular. Assim, as regiões funcionalmente significativas de fibrilas de colágeno que são fisicamente protegidas dentro da fibrila podem ser preferencialmente preservadas em fósseis.
Esses resultados mostram empiricamente que as relações estrutura-função a nível molecular podem contribuir para a preservação seletiva em restos de vertebrados fossilizados ao longo do tempo geológico, sugerem um "motivo de preservação" e reforçam os conceitos atuais que ligam a estrutura do colágeno à função biológica. Esta distribuição não aleatória suporta a hipótese de que os peptídeos são produzidos pelos organismos extintos e sugere um mecanismo químico para a sobrevivência.
San Antonio et al. [14] ressaltaram que as extrapolações de estudos [in vitro] da degradação de proteínas em condições aceleradas de acidez e alta temperatura prevêem que os fios de proteína não podem durar mais de alguns milhões de anos (a 10 ° C), mas sugerem que esses modelos podem não ser apropriados, já que "eles não consideram as moléculas em seu estado nativo (ou seja, dobradas, bem embaladas, reticuladas ou, no caso dos ossos, estabilizadas por associação com a fase mineral) ".
Demarchi et al. [31] recentemente quantificaram e descreveram mecanicamente a estabilização de proteínas que estão ligadas a uma superfície mineral. Eles descobriram que esse efeito de superfície explica a sobrevivência de proteínas originais em conchas de ovos de avestruz fóssil em climas africanos quentes durante pelo menos 3,8 milhões de anos, o que é mais longo do que o esperado. Eles calcularam que isso corresponde à sobrevivência da proteína por pelo menos 16 milhões de anos a uma temperatura constante mais fria de 10ºC, como seria típico do noroeste da Europa.
Fazale Rana da Reasons to Believe (um ministério cristão evangélico que aceita uma terra velha) desembalou o significado dessas descobertas [15]:
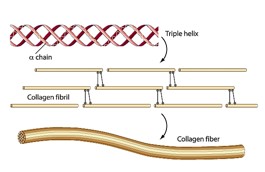
A unidade estrutural básica do colágeno é chamada de hélice tripla, constituída por três cadeias de proteína que se entrelaçam uma em torno da outra. Em certos pontos ao longo da hélice tripla, as cadeias de proteínas individuais são quimicamente ligadas entre si para formar reticulados.
Numerosas hélices triplas de colágeno se agrupam de forma escalonada para formar uma estrutura maior chamada fibrila de colágeno. Um grande número de fibrilas de colágeno, por sua vez, montam, com a ajuda de outras proteínas, fibras de colágeno.
Fazale Rana da Reasons to Believe (um ministério cristão evangélico que aceita uma terra velha) desembalou o significado dessas descobertas [15]:
A unidade estrutural básica do colágeno é chamada de hélice tripla, constituída por três cadeias de proteína que se entrelaçam uma em torno da outra. Em certos pontos ao longo da hélice tripla, as cadeias de proteínas individuais são quimicamente ligadas entre si para formar reticulados.
Numerosas hélices triplas de colágeno se agrupam de forma escalonada para formar uma estrutura maior chamada fibrila de colágeno. Um grande número de fibrilas de colágeno, por sua vez, montam, com a ajuda de outras proteínas, fibras de colágeno.
 |
| Fonte: Fazale Rana, “Structure of Dinosaur Collagen Unravels the Case for a Young Earth”, Reasons to Believe, August 10, 2011. |
A estrutura de colágeno altamente entrelaçada e reticulada torna razoável que fragmentos desta molécula possam sobreviver por 68 milhões de anos. Mesmo que as fibras de proteína individuais se quebram, a fibra ainda permaneceria intacta por causa de todos os pontos de associação. Uma vez que a cadeia de proteína se quebra, os fragmentos são mantidos em proximidade dos pontos de contato. Essa proximidade forçada permite que os fios quebrados ocasionalmente se reúnam e reformem a proteína original. Se os fios quebrados não se mantinham justapostos uns aos outros, os fragmentos se difundiriam um para o outro, impedindo assim a reversão do processo de degradação.
Finalmente, a associação do colágeno com a matriz óssea proporciona maior estabilidade às proteínas do colágeno. Dentro da matriz óssea, as fibras de colágeno adsorvem ao componente mineral do osso. O contato com a superfície protege a proteína e mantém os pedaços de colágeno justapostos sempre que as fibras de proteínas se quebram.
Finalmente, a associação do colágeno com a matriz óssea proporciona maior estabilidade às proteínas do colágeno. Dentro da matriz óssea, as fibras de colágeno adsorvem ao componente mineral do osso. O contato com a superfície protege a proteína e mantém os pedaços de colágeno justapostos sempre que as fibras de proteínas se quebram.
Em 2016, Rana, bioquímica por profissão, publicou o Dinosaur Blood e and the Age of the Earth, um pequeno livro dedicado às questões sobre os tecidos moles de dinossauro. Depois de descrever achados de tecido mole em dinossauro e outros fósseis antigos, ele explica por que eles não significam que a terra é jovem. Ele desmascara o criacionista da terra jovem que afirma que a datação radiogênica das rochas não é confiável e descreve nove condições ou mecanismos que podem funcionar em conjunto para ajudar a preservar remanescentes de proteína em ossos de dinossauro.
Após a morte, as enzimas destruidoras de células podem ser soltas no corpo, acelerando a decomposição. No entanto, em uma revisão de 2002 sobre a sobrevivência da matéria orgânica no osso, Collins et al. [16] afirmou que "o colágeno no osso é protegido pela exclusão física de enzimas extracelulares microbianas". Segundo Peterson et al. [17], depois que os micróbios invadem os poros mais externos dos ossos de dinossauro, suas atividades metabólicas levam a depósitos minerais que selam hermeticamente as zonas mais internas dos ossos contra ataques de micróbios.
Após a morte, as enzimas destruidoras de células podem ser soltas no corpo, acelerando a decomposição. No entanto, em uma revisão de 2002 sobre a sobrevivência da matéria orgânica no osso, Collins et al. [16] afirmou que "o colágeno no osso é protegido pela exclusão física de enzimas extracelulares microbianas". Segundo Peterson et al. [17], depois que os micróbios invadem os poros mais externos dos ossos de dinossauro, suas atividades metabólicas levam a depósitos minerais que selam hermeticamente as zonas mais internas dos ossos contra ataques de micróbios.
Osteócitos, traços de DNA e ferro como conservante
Um artigo de 2013 no Bone by Schweitzer et al. [18] abrangeu uma série de tópicos. Enquanto seus estudos anteriores tinham tratado principalmente da caracterização de estruturas baseadas em colágeno em ossos de dinossauro, aqui ela se concentrou no que parecia ser os remanescentes de osteócitos. Osteócitos são o tipo mais comum de células encontradas no osso. Eles ajudam a regular a química da mineralização do osso, para manter a massa óssea apropriada. Para obter os restos de osteócitos, ela primeiro removeu a porção mineral de ossos de dois dinossauros usando soluções de ácido fraco, conforme descrito anteriormente. Então, para libertar os osteócitos de sua matriz de colágeno, ela usou uma enzima que decompõe colágeno. Os osteócitos recuperados eram avermelhados, devido a um alto teor de ferro e assemelhavam-se fisicamente a osteócitos extraídos de aves modernas.
Schweitzer et al. [18] utilizaram anticorpos que se ligam a proteínas específicas para demonstrar a presença de quatro proteínas que se espera que estejam nos osteócitos, a saber, a actina, a tubulina, o PHEX e a histona H4. Os padrões de ligação desses anticorpos nos osteócitos de dinossauro corresponderam aos padrões vistos para ligação a osteócitos extraídos de avestruz. Os anticorpos não se ligaram ao tecido da matriz circundante ou a sedimentos associados. A espectrometria de massa também encontrou sequências de aminoácidos consistentes com essas quatro proteínas. A actina, a tubulina e a histona H4 não são encontradas nas bactérias, por isso é uma evidência adicional contra a explicação do biofilme desses tecidos moles.
Um artigo de 2013 no Bone by Schweitzer et al. [18] abrangeu uma série de tópicos. Enquanto seus estudos anteriores tinham tratado principalmente da caracterização de estruturas baseadas em colágeno em ossos de dinossauro, aqui ela se concentrou no que parecia ser os remanescentes de osteócitos. Osteócitos são o tipo mais comum de células encontradas no osso. Eles ajudam a regular a química da mineralização do osso, para manter a massa óssea apropriada. Para obter os restos de osteócitos, ela primeiro removeu a porção mineral de ossos de dois dinossauros usando soluções de ácido fraco, conforme descrito anteriormente. Então, para libertar os osteócitos de sua matriz de colágeno, ela usou uma enzima que decompõe colágeno. Os osteócitos recuperados eram avermelhados, devido a um alto teor de ferro e assemelhavam-se fisicamente a osteócitos extraídos de aves modernas.
Schweitzer et al. [18] utilizaram anticorpos que se ligam a proteínas específicas para demonstrar a presença de quatro proteínas que se espera que estejam nos osteócitos, a saber, a actina, a tubulina, o PHEX e a histona H4. Os padrões de ligação desses anticorpos nos osteócitos de dinossauro corresponderam aos padrões vistos para ligação a osteócitos extraídos de avestruz. Os anticorpos não se ligaram ao tecido da matriz circundante ou a sedimentos associados. A espectrometria de massa também encontrou sequências de aminoácidos consistentes com essas quatro proteínas. A actina, a tubulina e a histona H4 não são encontradas nas bactérias, por isso é uma evidência adicional contra a explicação do biofilme desses tecidos moles.
Um anticorpo PHEX específico, chamado OB 7.3, foi usado aqui. Entre os táxons existentes testados, ele se liga somente aqueles das aves. Foi mostrado neste estudo que se liga fortemente aos osteócitos de dinossauro e avestruz, mas não ao osteócito de jacaré.
As proteínas histonas, como a H4 encontrada nos ossos do dinossauro, estão associadas ao DNA. A presença de fragmentos de DNA real nos osteócitos de dinossauro é sugerida pela ligação de um anticorpo para o esqueleto do DNA e pela ligação de duas manchas químicas conhecidas como PI e DAPI. A coloração das células de dinossauro mostrou padrões espaciais similares às manchas em osteócitos de avestruz, apoiando a visão de que esse DNA é de células originais de dinossauro e não de infiltração microbiana. Essas manchas podem se ligar a cadeias de DNA tão baixas quanto 4 a 20 pares de bases, portanto, esses resultados não exigem que se tenham preservado sequências longas. Apenas cerca de 15-20% das células de dinossauro mostraram qualquer resposta à coloração, e para aqueles que reagiram, a coloração era muito menos intensa do que para o avestruz. Assim, quaisquer remanescentes de DNA nos ossos de dinossauro são bastante degradados. A equipe de Schweitzer só pode detectar vestígios de DNA nus, o que não é o suficiente para fazer sequenciação, de modo a verificar que este seja realmente DNA de dinossauro: "Esses dados não são suficientes para sustentar a afirmação de que o DNA visualizado nessas células é de origem dinossauro".
As proteínas histonas, como a H4 encontrada nos ossos do dinossauro, estão associadas ao DNA. A presença de fragmentos de DNA real nos osteócitos de dinossauro é sugerida pela ligação de um anticorpo para o esqueleto do DNA e pela ligação de duas manchas químicas conhecidas como PI e DAPI. A coloração das células de dinossauro mostrou padrões espaciais similares às manchas em osteócitos de avestruz, apoiando a visão de que esse DNA é de células originais de dinossauro e não de infiltração microbiana. Essas manchas podem se ligar a cadeias de DNA tão baixas quanto 4 a 20 pares de bases, portanto, esses resultados não exigem que se tenham preservado sequências longas. Apenas cerca de 15-20% das células de dinossauro mostraram qualquer resposta à coloração, e para aqueles que reagiram, a coloração era muito menos intensa do que para o avestruz. Assim, quaisquer remanescentes de DNA nos ossos de dinossauro são bastante degradados. A equipe de Schweitzer só pode detectar vestígios de DNA nus, o que não é o suficiente para fazer sequenciação, de modo a verificar que este seja realmente DNA de dinossauro: "Esses dados não são suficientes para sustentar a afirmação de que o DNA visualizado nessas células é de origem dinossauro".
Em uma entrevista em 2016 [29], Schweitzer observou:
"Encontrei DNA em osso de dinossauro", disse Mary Schweitzer, paleontóloga molecular da North Carolina State University. "Mas não o sequênciamos, não conseguimos recuperá-lo, e não podemos caracterizá-lo. A quem pertence é um mistério ".
Não é surpresa que o dinossauro contenha DNA, disse ela. Os ossos são parcialmente constituídos por um mineral chamado hidroxiapatita, que tem uma forte afinidade por certas biomoléculas, incluindo DNA. Na verdade, os pesquisadores geralmente usam hidroxiapatita para purificar e concentrar DNA no laboratório, disse Schweitzer.
"Essa é uma das razões pelas quais eu não trabalho com o DNA", disse Schweitzer à Live Science. "É muito propenso à contaminação e é realmente difícil de interpretar".
A pobre mostra de DNA nos fósseis de dinossauro é uma pancada no criacionismo da Terra jovem: se os dinossauros fossem enterrados em uma grande inundação apenas 4500 anos atrás, deveríamos encontrar gotas de DNA com longas sequências em pelo menos alguns dos restos de dinossauros em nossa posse. Agora é comum a sequência de DNA de certos outros achados fósseis, mas não de dinossauros.
"Encontrei DNA em osso de dinossauro", disse Mary Schweitzer, paleontóloga molecular da North Carolina State University. "Mas não o sequênciamos, não conseguimos recuperá-lo, e não podemos caracterizá-lo. A quem pertence é um mistério ".
Não é surpresa que o dinossauro contenha DNA, disse ela. Os ossos são parcialmente constituídos por um mineral chamado hidroxiapatita, que tem uma forte afinidade por certas biomoléculas, incluindo DNA. Na verdade, os pesquisadores geralmente usam hidroxiapatita para purificar e concentrar DNA no laboratório, disse Schweitzer.
"Essa é uma das razões pelas quais eu não trabalho com o DNA", disse Schweitzer à Live Science. "É muito propenso à contaminação e é realmente difícil de interpretar".
A pobre mostra de DNA nos fósseis de dinossauro é uma pancada no criacionismo da Terra jovem: se os dinossauros fossem enterrados em uma grande inundação apenas 4500 anos atrás, deveríamos encontrar gotas de DNA com longas sequências em pelo menos alguns dos restos de dinossauros em nossa posse. Agora é comum a sequência de DNA de certos outros achados fósseis, mas não de dinossauros.
Em 2012, Allentoft et al. [19] publicaram um estudo do DNA preservado em ossos de espécimes de moa que foram escavados na Nova Zelândia a partir de sedimentos anóxicos, tamponados com calcário. O moa era uma grande ave que não voava e era o herbívoro dominante na Nova Zelândia antes que os Maori chegassem e os perseguisse até a extinção. Neste estudo, os ossos da perna esquerda de 158 esqueletos de moa foram datados por radiocarbono e avaliados quanto ao conteúdo de DNA. As datas para esses esqueletos variaram de 602 a 7839 anos antes do presente, com todos os espécimes, exceto um, mais jovens do que cerca de 5800 anos. Os autores propuseram um ajuste exponencial aos dados, com uma meia vida de 521 anos para DNA mitocondrial. Este modelo prediz que o DNA se degradaria completamente para um único par de bases em 131.000 anos a 15 ° C (59 ° F), em 882.000 anos a 5 ° C (41 ° F) e em 6.830.000 anos a uma temperatura glacial de -5 ° C (23 ° F ).
Isso parece impedir a sobrevivência de qualquer fragmento de DNA nos ossos de dinossauro de 68 milhões de anos no estudo de Schweitzer, et al. [18]. No entanto, os dados de Allentoft et al. [19] estavam altamente espalhados; apenas 38,6% da variação na degradação do DNA entre amostras de osso de moa foi explicada pelas diferenças de idade. Para o conjunto de resultados que todos datam de cerca de 2700 anos atrás, os valores de DNA variam de cerca de 0,002 a 100. Esse é um fator de diferença de 50,000 do menor para o maior, mostrando a enorme variabilidade de resultados apenas dentro desse relativamente pequeno e homogêneo conjunto de dados.
Isso indica que a taxa de decaimento do DNA pode variar amplamente de amostra para amostra devido a vários fatores ambientais. Por exemplo, as taxas de degradação do DNA nos ossos moa foram quase 400 vezes mais lentas do que as previsões dos dados cinéticos publicados da decomposição in vitro do DNA. Assim, não é claro que estes resultados relativos a curto prazo (com menos de 8000 anos de idade) com ossos de pássaro em um cenário de calcário necessariamente prevejam o destino do DNA em outros espécimes, como os ossos de dinossauro muito maiores que foram depositados em arenito. Como outro exemplo da longevidade do DNA em algumas condições, fragmentos de DNA mitocondrial, suficientemente longos para sequenciar, foram recentemente recuperados de um fígado de hominínio de 400.000 anos de idade [20].
Isso indica que a taxa de decaimento do DNA pode variar amplamente de amostra para amostra devido a vários fatores ambientais. Por exemplo, as taxas de degradação do DNA nos ossos moa foram quase 400 vezes mais lentas do que as previsões dos dados cinéticos publicados da decomposição in vitro do DNA. Assim, não é claro que estes resultados relativos a curto prazo (com menos de 8000 anos de idade) com ossos de pássaro em um cenário de calcário necessariamente prevejam o destino do DNA em outros espécimes, como os ossos de dinossauro muito maiores que foram depositados em arenito. Como outro exemplo da longevidade do DNA em algumas condições, fragmentos de DNA mitocondrial, suficientemente longos para sequenciar, foram recentemente recuperados de um fígado de hominínio de 400.000 anos de idade [20].
Schweitzer et al. [18] propuseram uma série de mecanismos moleculares para a preservação a longo prazo das células e alguns de seus componentes químicos dentro dos ossos de dinossauro. Esses fatores incluem o isolamento dentro de pequenos poros no osso, associação com o mineral ósseo, as estruturas terciárias das proteínas e o papel do ferro na promoção da reticulação. Vou citar toda essa seção desse artigo, como de costume, omitido as referências bibliográficas:
As células geralmente são completamente degradadas logo após a morte do organismo, então como as células e as moléculas que as compõem podem persistir em osso mesozoico? Na matriz mineralizada do osso, muitos fatores convergem para alterar a dinâmica da morte celular e degradação, contribuindo para a interrupção da via de degradação. Por exemplo, as células necróticas ou apoptóticas são rapidamente destruídas por fagocitose ou por ataque microbiano pós-mortem, mas osteócitos são inacessíveis a outras células vivas, o que pode, em parte, explicar sua preservação nesses tecidos antigos. Em segundo lugar, os osteócitos são inerentemente resistentes à degradação porque a localização dentro da matriz óssea inibe a divisão celular, portanto, células podem ser necessárias para durar a vida útil do organismo. A expressão em osteócitos das proteínas repressoras apoptóticas também pode contribuir para a sua persistência. A associação de actina com alfa-actina e fimbrina confere estabilidade à actina ao longo da vida da célula e também pode estabilizar a proteína após a morte. Finalmente, osteócitos têm acesso limitado ao oxigênio dentro da matriz óssea e, portanto, podem ser protegidos contra danos oxidativos.
A morte celular, seja por apoptose ou necrose, é rapidamente seguida por autólise, que normalmente destrói a célula e libera enzimas autolíticas no ambiente circundante. A autólise, no entanto, é auto-limitante, e depois de atingir um determinado limite, as células restantes são estáveis por longos períodos.
A associação com mineral proporciona outras proteções que não estão disponíveis para tecidos e células não biomineralizados. As superfícies microcristalinas da apatite podem atuar como grãos de argila, adsorvendo enzimas degradativas e inativando-as, e além de limitar o acesso de micróbios a osteócitos, a matriz óssea rígida também pode inibir a desnaturação e o inchaço molecular que precede a autólise.
A morte celular, seja por apoptose ou necrose, é rapidamente seguida por autólise, que normalmente destrói a célula e libera enzimas autolíticas no ambiente circundante. A autólise, no entanto, é auto-limitante, e depois de atingir um determinado limite, as células restantes são estáveis por longos períodos.
A associação com mineral proporciona outras proteções que não estão disponíveis para tecidos e células não biomineralizados. As superfícies microcristalinas da apatite podem atuar como grãos de argila, adsorvendo enzimas degradativas e inativando-as, e além de limitar o acesso de micróbios a osteócitos, a matriz óssea rígida também pode inibir a desnaturação e o inchaço molecular que precede a autólise.
Em 2007, hipotetizamos que o ferro, liberado após a morte de hemoglobina e mioglobina através de autólise/degradação de glóbulos vermelhos e tecido muscular, atuaria para "fixar" tanto os tecidos quanto as moléculas, uma hipótese também apresentada por outros Fe (II) biologicamente ativos, que é solúvel, converteria rapidamente em Fe (III) após a libertação da célula e precipitaria da solução. O ferro é uma espécie reativa de oxigênio (ROS), e este interruptor desencadeia a formação de radicais hidroxílicos. Através de uma cascata de eventos referidos como Química de Fenton, os radicais hidroxila altamente reativos desencadeiam a reticulação de proteínas e a peroxidação e reticulação dos ácidos graxos que compõem as membranas celulares. Como os osteócitos estão intimamente ligados através das filopodias ao sistema vascular do osso e, como a ferritina da proteína de ligação ao ferro foi identificada nesta linha celular, eles seriam suscetíveis a essa reação em cadeia.
O ferro está implicado na preservação dessas microestruturas transparentes por sua associação íntima com as "membranas" celulares na TEM analítica (Fig. S1), e ainda apoiada pelo aumento da resposta de anticorpos em todos os casos em que essas "células" de dinossauro foram tratadas com o quelador de ferro PIH (ver Métodos experimentais ampliados no apêndice A). Se as membranas dessas "células" fossem preservadas através de reticulação mediada pelo ferro (Fig S1), ela também fornece um mecanismo pelo qual as proteínas associadas à membrana podem ser preservadas. A actina e a tubulina têm uma estrutura terciária que os torna inerentemente resistentes à degradação precoce, mas sua estreita associação com uma membrana naturalmente "fixa" aumentaria muito o seu potencial de preservação. Da mesma forma, PHEX é uma proteína transmembranar, e as regiões da proteína incorporadas dentro da membrana seriam "fixadas" naturalmente junto com a membrana. O ferro também pode funcionar para ligar o oxigênio, evitando danos oxidativos aos tecidos e moléculas. Finalmente, uma vez que as "células" são fixadas desta maneira, elas podem ser ainda mais estabilizadas por mobilização e precipitação mediada por modelo de apatita microcristalina no início da diagênese.
Em outro estudo de 2013, Schweitzer et al. [21] examinaram experimentalmente o papel do ferro na preservação das proteínas. Depois que um animal morre, o ferro da hemoglobina nos glóbulos vermelhos pode ser liberado para interagir com outros tecidos. Citam publicações anteriores que descrevem como o ferro pode facilitar a formação de radicais oxi, que "facilitam a reticulação proteica de maneira análoga às ações de fixadores de tecidos (por exemplo, formaldeído), aumentando assim a resistência dessas biomoléculas "fixadas" a enzimas ou digestão microbiana". Usando uma série de técnicas analíticas, eles observaram nanopartículas ricas em ferro como sendo intimamente associadas ao tecido vascular flexível preservado recuperado dos ossos de T. rex e dinossauros bico-de-pato. A imagem de alta ampliação abaixo mostra nanopartículas inorgânicas ricas em ferro associadas à camada orgânica do vaso de um T. rex. Nesta imagem, as partículas de ferro parecem estar concentradas no exterior do recipiente. O tecido do vaso bico-de-pato (ver Figura 1 (b) de Schweitzer et al. [21]) é mais uniformemente infiltrado por partículas de ferro. Parece-me provável que o ferro fosse originalmente mais finamente disperso e mais disponível para fazer química e depois precipitou nessas nanopartículas.
O contato direto com ferro ou partículas de ferro, como no contato direto com mineral ósseo, pode auxiliar na estabilização do tecido. Nessas imagens, características celulares internas, como a cromatina e a membrana nuclear, eram visíveis no tecido de avestruz, mas não nas estruturas de dinossauro, o que é consistente com a degradação substancial de natureza interna das células do dinossauro.
![Transmission Electron Microscope (TEM) image of vessel from T. rex bone, showing iron-rich nanoparticles associated with organic layer. Source: Schweitzer, et al., Proc. R. Soc. B 281: 20132741 [21].](https://letterstocreationists.files.wordpress.com/2015/02/tem-of-vessel-with-iron-particles.jpg?w=640) |
| Imagem do microscópio eletrônico de transmissão (TEM) de vaso do osso de T. rex, mostrando nanopartículas ricas em ferro associadas à camada orgânica. Fonte: Schweitzer, et al., Proc. R. Soc. B 281: 20132741 [21]. |
Quando o ferro foi quimicamente removido dos vasos por tratamento com agentes quelantes, a resposta dos tecidos do vaso a anticorpos proteicos específicos aumentou dramaticamente. Esta é outra indicação da associação do ferro residual com estas proteínas preservadas.
Schweitzer et al. [21] também incubaram vasos sanguíneos de avestruzes em uma variedade de soluções para testar o efeito do ferro derivado da hemoglobina na preservação do tecido. Alguns vasos de avestruz foram incubados em uma solução de hemoglobina. Esta hemoglobina foi extraída dos glóbulos vermelhos do sangue de galinha e avestruz e depois foi diluída novamente para a concentração original no sangue aviano. Os vasos assentados na solução de hemoglobina não mostraram sinais de degradação por mais de dois anos. Em contrapartida, os vasos de avestruz em água potável ou solução salina tamponada com fosfato (PBS) apresentaram degradação significativa dentro de três dias, ou seja, mais de 240 vezes mais rápido de degradação do que com a hemoglobina. A figura abaixo compara o estado dos vasos sanguíneos após 30 dias de incubação com hemoglobina no meio de incubação (vasos quase todos intactos) ou sem hemoglobina (vasos quase todos decompostos). Esses resultados demonstram dramaticamente a eficácia da preservação do tecido à base de ferro, que geralmente não foi levada em consideração nas estimativas anteriores de quanto tempo as proteínas podem sobreviver.
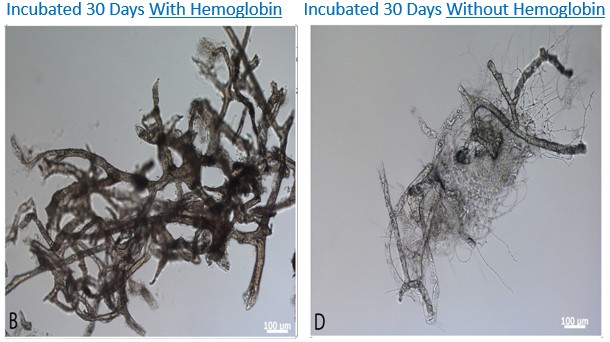 |
| Os vasos sanguíneos de avestruz incubaram-se durante 30 dias à temperatura ambiente sob condições oxigenadas, com (B) ou sem (D) adição de na solução de incubação. Da Figura S5 de Schweitzer et al. "A role for iron and oxygen chemistry in preserving soft tissues, cells and molecules from deep time", Proc. R. Soc. B 281: 20132741.http://rspb.royalsocietypublishing.org/content/royprsb/281/1775/20132741.full.pdf |
Um estudo de 2014 de Boatman et al. [22] envolveu mais experimentos para investigar efeito baseado em ferro nas proteínas. A espectroscopia infravermelha inicial sugeriu a presença de colágeno altamente reticulado no tecido fóssil de dinossauro. Os pesquisadores identificaram dois mecanismos prováveis de reticulação não enzimática, a reação e a glicação de Fenton. Ambas as reações dependem do potencial de oxidação do ferro. Para testar os papéis desses mecanismos, eles incubaram osso de frango fresco e desmineralizado usando tratamentos correspondentes para induzir reticulação de colágeno. Os resultados analíticos mostraram que esses tratamentos, de fato, induzem o tipo de reticulação que deverá tornar o colágeno mais resistente à decomposição. Além disso, os tecidos de dinossauro preservados foram suficientemente reticulados para resistir a um produto químico que corta as moléculas levemente reticuladas:
Os tecidos de frango desmineralizados incubados de acordo com a reação e glicação de Fenton produziram produtos consistentes com hiperreticulamento induzido ... Ambos os tecidos de frango tratados desenvolveram pequenos picos de ésteres (aproximadamente 1732 cm-1); em comparação, um pico de éster proeminente foi observado em todos os espectros de FTIR coletados para Tyrannosaurus rex. Os vasos fósseis tratados com o agente redutor [NaBH4, que pode clivar reticulações intermoleculares de baixa ordem] não produziram alterações significativas na análise FTIR, sugerindo que as reticulaçãoes não-enzimáticas formadas neste tecido são irredutíveis. Tais formações de ligação ocorrem entre três ou mais cadeias peptídicas e, como tal, tendem a ser altamente resistentes à clivagem redutora.
Avaliação da evidência de que o tecido mole pode persistir por 70 milhões de anos
Como seria de esperar, os criacionistas da terra jovem tomaram essas observações de tecidos moles de ossos de dinossauro como evidência de que esses fósseis não podem ter mais que alguns poucos milhares de anos – e, portanto, os métodos geológicos convencionais como a datação radioativa devem ser terrivelmente falhos, já que esses métodos mostra que as camadas das rochas que sepultaram esses fósseis têm cerca de 70 milhões de anos.
Como seria de esperar, os criacionistas da terra jovem tomaram essas observações de tecidos moles de ossos de dinossauro como evidência de que esses fósseis não podem ter mais que alguns poucos milhares de anos – e, portanto, os métodos geológicos convencionais como a datação radioativa devem ser terrivelmente falhos, já que esses métodos mostra que as camadas das rochas que sepultaram esses fósseis têm cerca de 70 milhões de anos.
Eu citei alguns desses artigos da terra jovem no início deste artigo. Googlar "dinosaur soft tissue age earth" produziu milhares de resultados. Dezenas e dezenas desses são sites que promovem o criacionismo da Terra jovem, alegando que os resultados de Schweitzer desprovam a evolução e a datação radioativa de rochas. Alguns desses sites deturpam os fatos, afirmando que os glóbulos vermelhos reais foram encontrados. Conforme mencionado acima, esse não é o caso: estas pequenas coisas vermelhas e redondas são pedaços de óxido de ferro, como a ferrugem, que acabam sendo do tamanhao e forma dos glóbulos vermelhos. Os restos orgânicos reais são remanescentes altamente reticulados de várias proteínas que são conhecidas por ter estruturas estáveis. Estes remanescentes mantêm a forma do tecido macio original, o que não é surpreendente, já que eles foram confinados dentro de pequenos poros nos ossos de dinossauro.
O principal ataque dos criacionistas da Terra jovem sobre a antiguidade desses achados é um argumento da incredulidade, baseado na ignorância: "É obviamente impossível que qualquer vestígio de tecido macio dure por 70 milhões de anos". Mas, como eles sabem que isso é impossível? Eles não sabem. Sim, experimentos sobre degradação de proteína em tubos de ensaio indicam que as proteínas se desintegrariam completamente em cerca de um milhão de anos. Mas muitos exemplos mostram que a taxa de degradação da proteína varia de forma selvagem, dependendo das condições, então ninguém pode dizer com certeza quanto tempo alguns fragmentos de proteína podem durar, preservados com ferro e selados em poros minerais. Nós simplesmente não sabemos como esse processo avança ao longo de um período de milhões de anos. É difícil inventar experimentos definitivos para simular esse período de tempo.
Os vasos de avestruz discutidos acima tiveram sua vida antes da degradação prolongada de três dias para mais de dois anos, um fator de mais de 240. Podemos até ver taxas de decomposição muito diferentes em nossos alimentos, dependendo de como é tratado. Se você deixar uma jarra de leite no balcão durante duas semanas, haverá muita bioquímica (principalmente fermentação de lactose em ácidos, mas também incluindo degradação de proteína) que pode torná-lo impróprio para consumo humano. No entanto, se o leite é contatado com as bactérias certas e outros materiais, as proteínas do leite podem ser preservadas sob a forma de queijo que pode ficar sentado na estante por anos. Da mesma forma, se a carne é moída para fazer hambúrguer e deixada em um pacote no balcão por uma semana, ela irá apodrecer. No entanto, se a carne for cortada em fatias finas e secadas, ela durará meses. O mesmo leite e a mesma carne, mas com um conjunto diferente de condições, eles podem reter proteínas por dez ou cem vezes mais, dependendo de suas circunstâncias.
Para outro exemplo, em estudos forenses na "fazenda corporal" da Universidade do Tennessee, os cadáveres doados são deliberadamente colocados sobre ou no chão para decadência natural para que os pesquisadores possam rastrear exatamente como eles se decompõem à medida que passam por etapas de inchaço, consumo por larvas, etc. De estudos como estes, sabemos que a carne e a pele de um humano podem decair em cerca de um mês em condições úmidas. Então, depois de um mês, o rosto de um homem pode parecer algo assim:
 |
| Fonte: http://www.shutterstock.com |
Agora, considere este indivíduo:
 |
| O homem de Tollund. Fonte: Wikipedia, “Tollund Man” |
Quando ele foi descoberto em uma turfa dinamarquesa, parecendo tão vivo, a polícia foi convocada com a suposição de que ele era uma vítima de assassinato recente. Este "Homem de Tollund" era de fato uma vítima de assassinato, mas o crime (provavelmente um sacrifício ritualístico humano) ocorreu há mais de 2200 anos. Acontece que as condições químicas no pântano em que ele foi jogado facilitaram a preservação da pele e alguns outros tecidos moles. Note-se que com esta e outras pessoas de pântano, a abordagem errada seria insistir, porque normalmente a pele humana não dura há milhares de anos, eles devem ter morrido recentemente. Em vez disso, os pesquisadores levaram em conta outras informações de datação para perceber que essas pessoas de pântano tinham mais de mil anos de idade, mesmo que o mecanismo de preservação não fosse inicialmente conhecido. Comparando 2200 anos de preservação aqui, versus a decomposição completa da carne em um mês do corpa da fazenda no Tennessee, temos um fator de diferença de mais de 25.000 nas taxas de degradação dos tecidos moles.
Isso novamente faz com que as taxas e os modos de decomposição de proteínas e tecidos moles possam variar drasticamente, dependendo das circunstâncias. Assim, é absurdo dizer que, porque as proteínas desaparecem em um milhão de anos sob um conjunto de condições, portanto, os remanescentes de proteína não podem suportar mais de 100 milhões de anos sob outras condições. A afirmação de que "sabemos que fragmentos substanciais de proteínas, mesmo em alguma forma reticulada, não podem sobreviver por 80 milhões de anos" simplesmente não é verdade. Uma vez que essa afirmação (de uma forma ou de outra) está no cerne da interpretação da Terra jovem desses tecidos fósseis, a defesea da Terra jovem colpasa aqui.
Continua aqui.
Referências
[11] Schweitzer, et al., Proc. R. Soc. B (2005) 272, 775–784 . http://rspb.royalsocietypublishing.org/content/royprsb/272/1565/775.full.pdf
[12] Mary Higby Schweitzer, “Soft Tissue Preservation in Terrestrial Mesozoic Vertebrates”, Annual Review of Earth and Planetary Sciences Vol. 39: 187-216 (May 2011) http://www.annualreviews.org/doi/abs/10.1146/annurev-earth-040610-133502
[13] Theagarten Lingham-Soliara and Joanna Glabb,” Dehydration: A
mechanism for the preservation of fine detail in fossilised soft tissue
of ancient terrestrial animals”, Palaeogeography, Palaeoclimatology, Palaeoecology, Volume 291, Issues 3–4, 15 May 2010, Pages 481–487 http://www.sciencedirect.com/science/article/pii/S0031018210001471
[14] James D. San Antonio, Mary H. Schweitzer, Shane T. Jensen,
Raghu Kalluri, Michael Buckley, Joseph P. R. O. Orgel. “Dinosaur
Peptides Suggest Mechanisms of Protein Survival”, PLoS One 6, no. 6 (2011): e20381 http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0020381
[15] Fazale Rana , “Structure of Dinosaur Collagen Unravels the Case for a Young Earth”, Reasons to Believe, August 10, 2011. http://www.reasons.org/articles/structure-of-dinosaur-collagen-unravels-the-case-for-a-young-earth
[16] M. J. Collins, C. M. Nielsen-Marsh, J. Hiller, C. I. Smith And
J. P. Roberts, “The Survival Of Organic Matter In Bone: A Review”, Archaeometry 44, 3 (2002) 383–39 http://onlinelibrary.wiley.com/doi/10.1111/1475-4754.t01-1-00071/pdf
[17] Peterson JE, Lenczewski ME, Scherer RP (2010) “Influence of
Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil
and Extant Archosaurs”. PLoS ONE 5(10): e13334. http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0013334
[18] Mary Higby Schweitzer, Wenxia Zheng, Timothy P. Cleland,
Marshall Bern. “Molecular analyses of dinosaur osteocytes support the
presence of endogenous molecules”. Bone Volume 52, Issue 1, January 2013, Pages 414–423. http://www4.ncsu.edu/~lezanno/Research_files/SchweitzerEtAl2012.pdf
[19] Morten E. Allentoft, et al.,”The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils”. Proc. R. Soc. B (2012) 279, 4724–4733 . http://rspb.royalsocietypublishing.org/content/279/1748/4724
[20] Matthias Meyer, et al., “A mitochondrial genome sequence of a hominin from Sima de los Huesos”, Nature 505, 403–406 . http://www.nature.com/nature/journal/v505/n7483/full/nature12788.html
[21] Schweitzer MH, Zheng W, Cleland TP, Goodwin MB, Boatman E,
Theil E, Marcus MA, Fakra SC. “A role for iron and oxygen chemistry in
preserving soft tissues, cells and molecules from deep time”, Proc. R.
Soc. B 281: 20132741. http://rspb.royalsocietypublishing.org/content/royprsb/281/1775/20132741.full.pdf
[22] Elizabeth M. Boatman, Mark B. Goodwin, Hoi-Ying Holman,
Sirine Fakra, Mary H. Schweitzer, Ronald Gronsky and John R. Horner,
“Synchrotron Chemical and Structural Analysis of Tyrannosaurus rex Blood Vessels:The Contribution of Collagen Hypercrosslinking to Tissue Longevity”, Microsc. Microanal. 20 (Suppl 3), 2014 http://infrared.als.lbl.gov/Publications/2014/BGHFSGH14/1430.pdf
[29] Laura Geggel, ” Is It Possible to Clone a Dinosaur?” LiveScience April 28, 2016 http://www.livescience.com/54574-can-we-clone-dinosaurs.html




Nenhum comentário:
Postar um comentário